
Heparan Sulfate Binding Cationic Peptides Restrict SARS-CoV-2 Entry
 , , and
, , and 
Abstract
:1. Introduction
2. Results
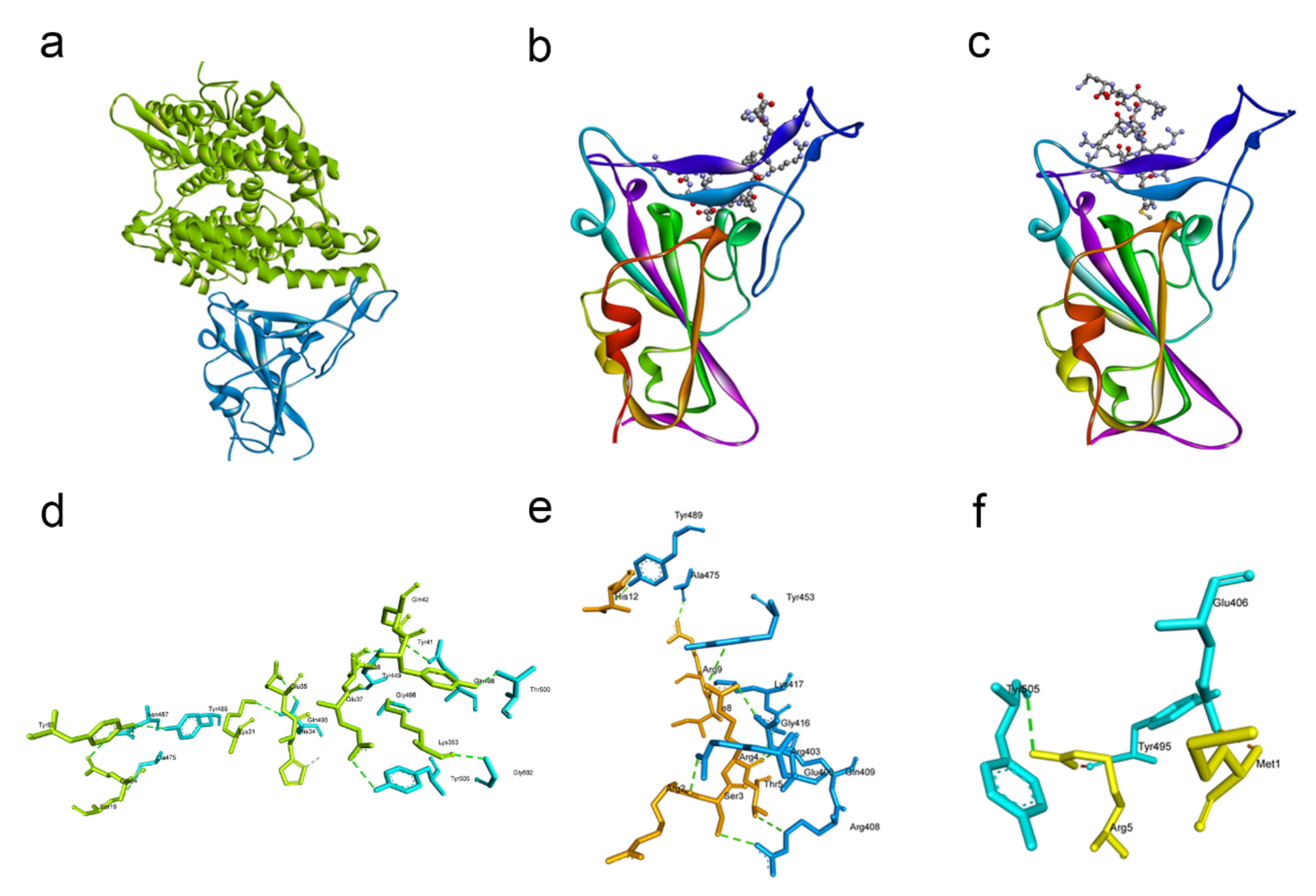
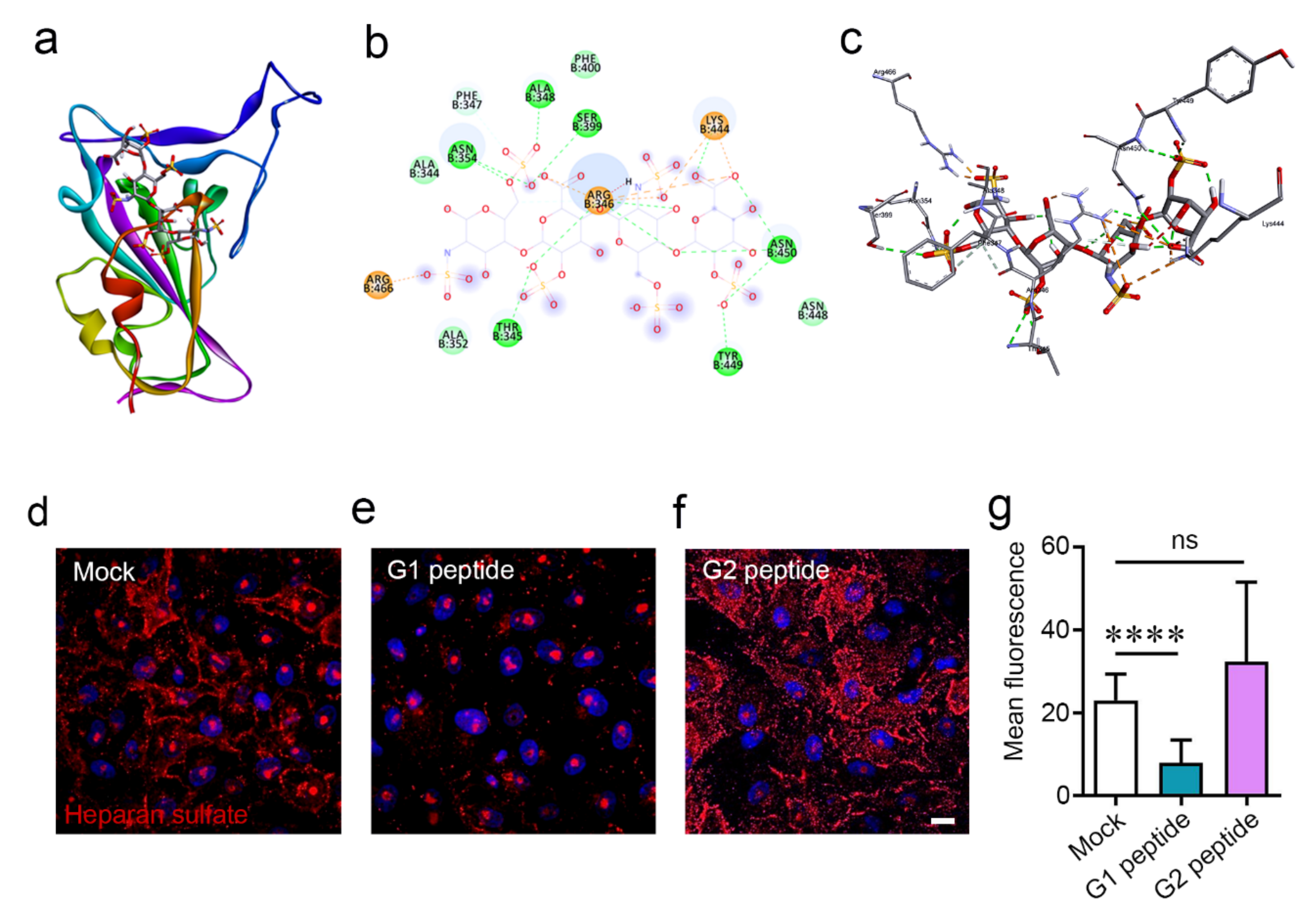
2.1. Docking Studies Revealed That G1 and G2 Peptides May Bind to SARS-CoV-2 and Inhibit Its Key Receptors
2.2. G1 and G2 Peptides Reduce Cell Surface Heparan Sulfate Expression
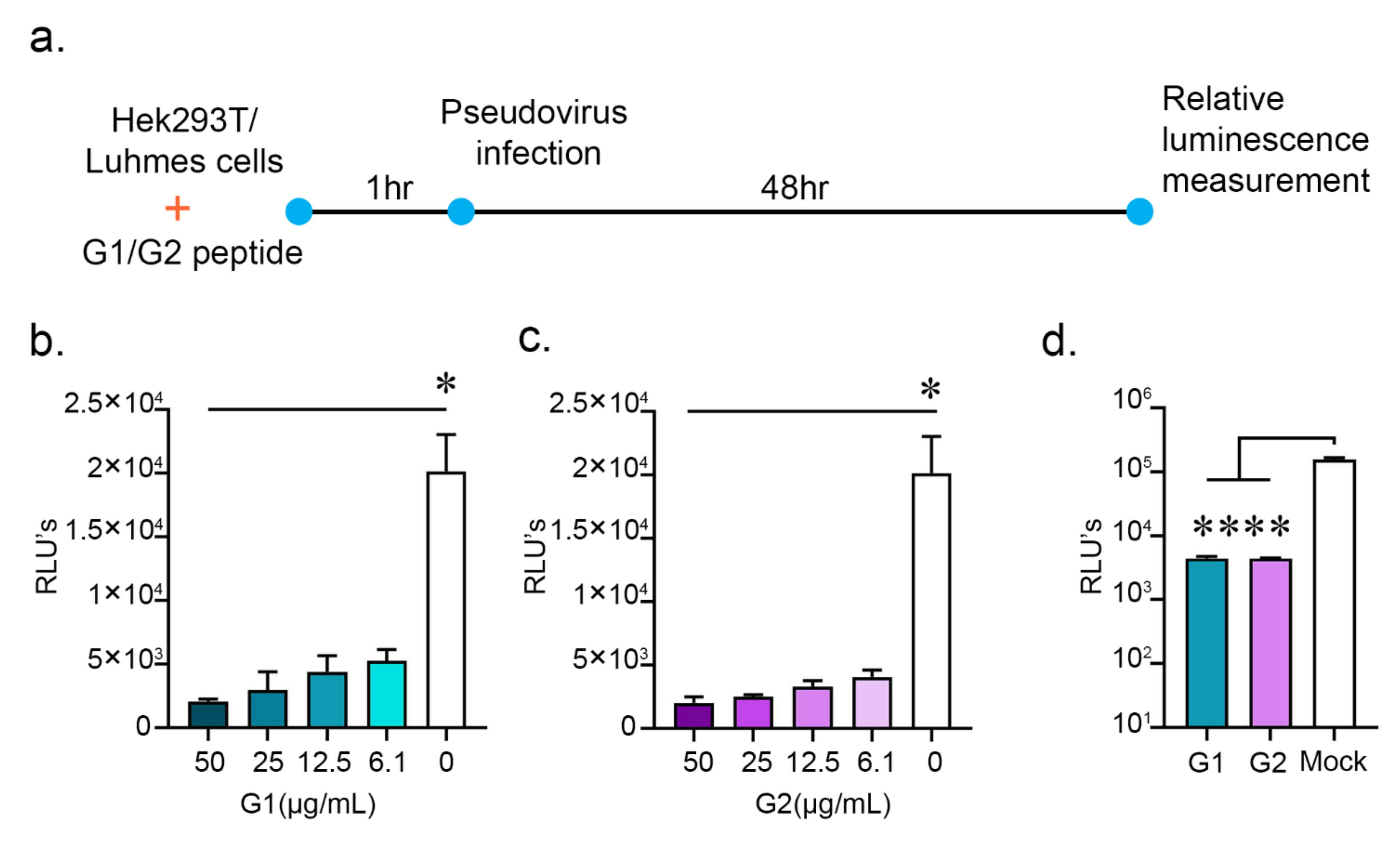
2.3. G1 and G2 Peptides Inhibit SARS-CoV-2 Pseudovirus Entry
3. Discussion
4. Materials and Methods
4.1. Cell Line
4.2. Peptide Synthesis and Preparation
4.3. Cell Viability (MTT) Assay
4.4. SARS-CoV-2 Pseudotyped Virus Particles Formation
4.5. Entry Assay
4.6. Heparan Sulfate Labelling
4.7. Homology Modeling
4.8. Docking Studies
4.9. Statistics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.; Wang, W.; Song, Z.; Hu, Y.; Tao, Z.; Tian, J.; Pei, Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadfield, J.; Bedford, T.; Neher, R.; Hodcroft, E.; Sibley, T.; Huddleston, J.; Aksamentov, I.; Lee, J.; Fay, K.; Zuber, M.; et al. Nextstrain SARS-CoV-2 Resources. Available online: https://nextstrain.org/sars-cov-2 (accessed on 20 March 2021).
- Johns Hopkins University. Mortality Analyses. Available online: https://coronavirus.jhu.edu/data/mortality (accessed on 20 March 2021).
- Clausen, T.M.; Sandoval, D.R.; Spliid, C.B.; Pihl, J.; Perrett, H.R.; Painter, C.D.; Narayanan, A.; Majowicz, S.A.; Kwong, E.M.; McVicar, R.N.; et al. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2. Cell 2020, 183, 1043–1057.e15. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chopra, P.; Li, X.; Wolfert, M.; Tompkins, M.; Boons, G. SARS-CoV-2 spike protein binds heparan sulfate in a length- and sequence-dependent manner. BioRxiv 2020. [Google Scholar] [CrossRef]
- Kwon, P.S.; Oh, H.; Kwon, S.; Jin, W.; Zhang, F.; Fraser, K.; Hong, J.J.; Linhardt, R.J.; Dordick, J.S. Sulfated polysaccharides effectively inhibit SARS-CoV-2 in vitro. Cell Discov. 2020, 6, 50. [Google Scholar] [CrossRef]
- Lang, J.; Yang, N.; Deng, J.; Liu, K.; Yang, P.; Zhang, G.; Jiang, C. Inhibition of SARS pseudovirus cell entry by lactoferrin binding to heparan sulfate proteoglycans. PLoS ONE 2011, 6, e23710. [Google Scholar] [CrossRef]
- Dogra, P.; Martin, E.B.; Williams, A.; Richardson, R.L.; Foster, J.S.; Hackenback, N.; Kennel, S.J.; Sparer, T.E.; Wall, J.S. Novel heparan sulfate-binding peptides for blocking herpesvirus entry. PLoS ONE 2015, 10, e0126239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, V.; Liu, J.; Valyi-Nagy, T.; Shukla, D. Anti-heparan sulfate peptides that block herpes simplex virus infection in vivo. J. Biol. Chem. 2011, 286, 25406–25415. [Google Scholar] [CrossRef] [Green Version]
- Jaishankar, D.; Buhrman, J.S.; Valyi-Nagy, T.; Gemeinhart, R.A.; Shukla, D. Extended release of an anti–heparan sulfate peptide from a contact lens suppresses corneal herpes simplex virus-1 infection. Invest. Ophthalmol. Vis. Sci. 2016, 57, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Koganti, R.; Suryawanshi, R.; Shukla, D. Heparanase, cell signaling, and viral infections. Cell Mol. Life Sci. 2020, 77, 5059–5077. [Google Scholar] [CrossRef]
- Stewart, M.D.; Sanderson, R.D. Heparan sulfate in the nucleus and its control of cellular functions. Matrix Biol. 2014, 35, 56–59. [Google Scholar] [CrossRef]
- Helms, J.; Kremer, S.; Merdji, H.; Clere-Jehl, R.; Schenck, M.; Kummerlen, C.; Collange, O.; Boulay, C.; Fafi-Kremer, S.; Ohana, M.; et al. Neurologic Features in Severe SARS-CoV-2 Infection. N. Engl. J. Med. 2020, 382, 2268–2270. [Google Scholar] [CrossRef]
- Zhang, B.; Chu, H.; Han, S.; Shuai, H.; Deng, J.; Hu, Y.; Gong, H.; Lee, A.C.; Zou, Z.; Yau, T.; et al. SARS-CoV-2 infects human neural progenitor cells and brain organoids. Cell Res. 2020, 30, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Ali, M.M.; Karasneh, G.A.; Jarding, M.J.; Tiwari, V.; Shukla, D. A 3-O-sulfated heparan sulfate binding peptide preferentially targets herpes simplex virus 2-infected cells. J. Virol. 2012, 86, 6434–6443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Koganti, R.; Lambe, U.P.; Yadavalli, T.; Nandi, S.S.; Shukla, D. Vaccines and Therapies in Development for SARS-CoV-2 Infections. J. Clin. Med. 2020, 9, 1885. [Google Scholar] [CrossRef]
- Edwards, T.G.; Bloom, D.C. Lund human mesencephalic (LUHMES) neuronal cell line supports herpes simplex virus 1 latency in vitro. J. Virol. 2019, 93, e02210-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millet, J.K.; Tang, T.; Nathan, L.; Jaimes, J.A.; Hsu, H.; Daniel, S.; Whittaker, G.R. Production of pseudotyped particles to study highly pathogenic coronaviruses in a biosafety level 2 setting. J. Vis. Exp. 2019, 145, 3791/59010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surya, W.; Li, Y.; Torres, J. Structural model of the SARS coronavirus E channel in LMPG micelles. Biochim. Biophys. Acta Biomembr. 2018, 1860, 1309–1317. [Google Scholar] [CrossRef]
- Ko, J.; Park, H.; Heo, L.; Seok, C. GalaxyWEB server for protein structure prediction and refinement. Nucleic Acids Res. 2012, 40, 294. [Google Scholar] [CrossRef]
- Davis, I.W.; Leaver-Fay, A.; Chen, V.B.; Block, J.N.; Kapral, G.J.; Wang, X.; Murray, L.W.; Arendall, W.B.; Snoeyink, J.; Richardson, J.S.; et al. MolProbity: All-atom contacts and structure validation for proteins and nucleic acids. Nucleic Acids Res. 2007, 35, 375. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, 407. [Google Scholar] [CrossRef] [Green Version]
- Sippl, M.J. Recognition of errors in three-dimensional structures of proteins. Proteins 1993, 17, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Jin, B.; Li, H.; Huang, S. HPEPDOCK: A web server for blind peptide-protein docking based on a hierarchical algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein-protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Spike RBD Residue Involved in the Hydrogen Bond | ACE2 Residue Involved in the Hydrogen Bond |
|---|---|
| GLN493 | LYS31 |
| THR500 | TYR41 |
| GLN498 | GLN42 |
| TYR449 | ASP38 |
| ASN487 | GLN24 |
| GLN493 | GLU35 |
| TYR505 | GLU37 |
| TYR453 | HIS34 |
| ALA475 | SER19 |
| ASN487 | TYR83 |
| TYR489 | TYR83 |
| GLY496 | LYS353 |
| GLY502 | LYS353 |
| Spike RBD Residue Involved in the Hydrogen Bond | G1 Peptide Residue Involved in the Hydrogen Bond |
|---|---|
| GLU406 | ARG4 |
| ARG403 | ARG2 |
| ARG408 | SER3 |
| LYS417 | ILE8 |
| TYR489 | HIS12 |
| ALA475 | ARG9 |
| ARG408 | THR5 |
| GLY416 | ARG4 |
| TYR453 | ARG4 |
| GLN409 | ARG4 |
| Spike RBD Residue Involved in the Hydrogen Bond | G2 Peptide Residue Involved in the Hydrogen Bond |
|---|---|
| GLU406 | MET1 |
| TYR505 | ARG5 |
| TYR495 | ARG5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suryawanshi, R.K.; Patil, C.D.; Koganti, R.; Singh, S.K.; Ames, J.M.; Shukla, D. Heparan Sulfate Binding Cationic Peptides Restrict SARS-CoV-2 Entry. Pathogens 2021, 10, 803. https://doi.org/10.3390/pathogens10070803
Suryawanshi RK, Patil CD, Koganti R, Singh SK, Ames JM, Shukla D. Heparan Sulfate Binding Cationic Peptides Restrict SARS-CoV-2 Entry. Pathogens. 2021; 10(7):803. https://doi.org/10.3390/pathogens10070803
Chicago/Turabian StyleSuryawanshi, Rahul K., Chandrashekhar D. Patil, Raghuram Koganti, Sudhanshu Kumar Singh, Joshua M. Ames, and Deepak Shukla. 2021. "Heparan Sulfate Binding Cationic Peptides Restrict SARS-CoV-2 Entry" Pathogens 10, no. 7: 803. https://doi.org/10.3390/pathogens10070803
APA StyleSuryawanshi, R. K., Patil, C. D., Koganti, R., Singh, S. K., Ames, J. M., & Shukla, D. (2021). Heparan Sulfate Binding Cationic Peptides Restrict SARS-CoV-2 Entry. Pathogens, 10(7), 803. https://doi.org/10.3390/pathogens10070803